Golfinhos e baleias atuais, junto com os peixes-boi e dugongos (Sirênios), são mamíferos completamente adaptados ao ambiente aquático. Suas atividades como alimentação, locomoção, descanso e reprodução dependem inteiramente desse ambiente. Eles não precisam, por exemplo, retornar à terra para executarem essas ações, diferentemente do que ocorre em outros grupos de mamíferos aquáticos, como leões marinhos, focas ou lontras.Mas você já pensou em como esse processo aconteceu?
A origem dos primeiros cetáceos (grupo que inclui golfinhos e baleias) se deu a partir de animais completamente terrestres. Formas extintas aparentadas aos artiodáctilos (grande grupo que inclui cabras, bois, camelos, hipopótamos, etc.) começaram essa jornada há cerca de 50 milhões de anos atrás. Eles se adaptaram, com o passar do tempo, às diversas peculiaridades do ambiente aquático, como a maior viscosidade, densidade, empuxo e pressão hidrostática. Entre as principais adaptações desenvolvidas pelos cetáceos, modificações anatômicas associadas à natação foram algumas das mais fundamentais para sua sobrevivência nesse “novo” ambiente.
Indohyus major, um animal extinto do Eoceno, terrestre e herbívoro, relacionado aos primeiros cetáceos. Arte de Nobu Tamura CC BY 3.0.
Nos cetáceos modernos, diversas características anatômicas e comportamentais permitem manobras na água e facilitam o deslocamento desses animais nos oceano, mares e rios. A evolução de algumas características pode ser rastreada nos fósseis. Porém, há uma falta considerável de informações sobre uma parte delas. Uma relação ainda pouco compreendida, por exemplo, é como se deu a divergência entre os golfinhos (odontocetos) e as baleias (misticetos).
Cynthiacetus (esquerda), um cetáceo extinto completamente aquático do fim do Eoceno, e Ambulocetus natans, uma forma semi-aquática de cetáceo do início do Eoceno. Foto de Jean-Pierre Dalbéra, CC BY 2.0.
Abundantes esqueletos de cetáceos do Eoceno ilustram a transição da vida semiaquática para a completamente aquática, incluindo o desenvolvimento de um corpo alongado, cilíndrico e com extremidades afiladas (corpo fusiforme). Fósseis do Eoceno também demonstram o gradual processo de redução das patas traseiras e a migração das narinas em direção ao topo da cabeça. Entretanto, há uma raridade excepcional de esqueletos de cetáceos em rochas do Oligoceno, o período geológico seguinte ao Eoceno, e isso tem dificultado muito os esforços para compreender a evolução da força de natação dos cetáceos. No Eoceno, a natação ainda era controlada parcialmente pelas patas traseiras, mas com o tempo ela passa a ser exercida exclusivamente pela cauda robusta.
Em 2020 o pesquisador Robert W. Boessenecker, junto com outros colegas, publicaram a descrição de uma nova espécie de um raro golfinho fóssil de grande porte, encontrado em estratos do Oligoceno do sul da Califórnia (E.U.A). Materiais deste animal já eram conhecidos desde o século 19, mas eram muito fragmentados, o que impedia que pesquisadores conhecessem melhor a espécie. Boessenecker e colegas descobriram, na década de 1990, um espécime surpreendentemente bem preservado, ainda que parcial, que permitiu não só batizarem adequadamente o animal (Ankylorhiza tiedemani), como também estudarem a evolução de algumas características transicionais pouco conhecidas dos cetáceos.
Ankylorhiza tiedemani possuía diversas características compartilhadas entre as baleias e golfinhos, o que deu aos cientistas pistas preciosas sobre a evolução destes grupos. O tamanho e outras características do corpo animal indicam que eleera um predador ativo, de natação rápida, que dominou as águas do seu tempo, ocupando um nicho semelhante aos das grandes orcas atuais.
Esqueleto de Ankylorhiza tiedemani (BOESSENECKER et al., 2020).
O primeiro material descrito para essa espécie foi um crânio muito incompleto recuperado por volta de 1880, que, na época, foi atribuído ao gênero Squalodon. Com a descoberta do esqueleto mais completo, descrito por Boessenecker e colegas em 2020, novas análises foram feitas e descobriu-se que, na verdade, o material pertencia a um novo gênero, batizado de Ankylorhiza. A. tiedemani é considerado, até o momento, o maior Odontoceto do Oligoceno, com aproximadamente 4,8m de comprimento, tamanho não superado até o Mioceno, quando aparecem no registro fossilífero os primeiros grandes cachalotes.
A. tiedemani possui o crânio e mandíbula robustos, com uma dentição simplificada quando comparada com os basilossaurídeos, grupo de cetáceos mais antigos, que tinham os dentes cheios de cristas e pequenas pontas acessórias. As características dentárias de A. tiedemani indicavam que ele se tratava de um caçador com elevada força de mordida, semelhante às encontradas nos primeiros cachalotes. Seus dentes da parte frontal são um mistério, pois possuem um ângulo estranho de inserção no crânio. Eles apontam para frente, o que indica que podem ter sido utilizados para competição entre indivíduos do mesmo sexo, como fazem as baleias-bicudas atuais, ou empregados na captura e abate de presas.
Baleia-bicuda (Ziphius cavirostris). Nos machos adultos podem ver-se dois dentes na ponta do maxilar inferior que estão orientados para a frente. Foto de Eveha CC BY 3.0.
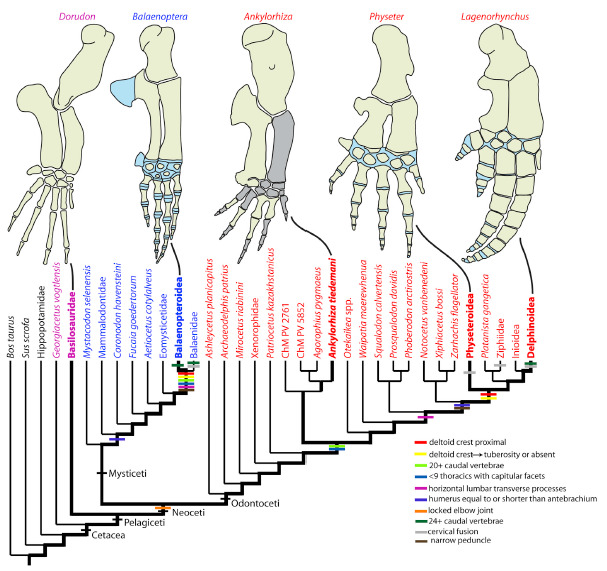
As nadadeiras peitorais de A. tiedemani possuem várias características derivadas, incluindo ossos longos (úmero, rádio e ulna) mais curtos quando comparados com os basilossaurídeos, porém, mais alongados quando comparados com os Odontocetos atuais. Suas nadadeiras e coluna vertebral também possuem características intermediárias, a maioria mais próxima de outros odontocetos basais, mas com algumas correlações com os misticetos. Isso coloca a espécie próxima à base da árvore evolutiva dos odontocetos.
Relações filogenéticas de Ankylorhiza tiedemani (BOESSENECKER et al., 2020).
A mobilidade de A. tiedemani seria semelhante à das falsas-orcas e orcas atuais, indicando uma natação reforçada, mais poderosa do que a dos basilossaurídeos (formas mais basais) de porte semelhante. Isso sugere que a espécie tinha velocidade suficiente para perseguir outros cetáceos,sirênios, tartarugas, aves marinhas, tubarões e outros peixes contemporâneos, incluindo esses organismos em sua dieta potencial.
A. tiedemani, finalmente, trouxe um pouco de luz sobre como diversas adaptações convergentes estavam presentes em odontocetos e misticetos basais, principalmente no que diz respeito a sua mobilidade. Futuras descobertas de espécimes mais completos ou ainda de novas espécies provenientes dos mesmos estratos geológicos podem ser chave na compreensão da evolução de mais aspectos da locomoção dos cetáceos modernos. Essas descobertas também podem auxiliar na elucidação de mais detalhes sobre como se deu a divergência entre baleias e golfinhos, um evento evolutivo fascinante e ainda pouco compreendido, que se deu nos mares do final do Eoceno e do início do Oligoceno.
Referência:
BOESSENECKER, Robert W. et al. Convergent evolution of swimming adaptations in modern whales revealed by a large macrophagous dolphin from the Oligocene of South Carolina. Current Biology, v. 30, n. 16, p. 3267-3273. e2, 2020.
Hoje (quarta-feira 9/12/2020), acaba de ser publicado um trabalho que traz informações importantes para a compreensão de um grande enigma da paleontologia: a origem dos pterossauros, répteis voadores que dominaram os céus durante a Era Mesozoica.
O trabalho foi publicado na revista Nature e conta com a participação de alguns cientistas brasileiros, que ajudaram a investigar em detalhes um grupo de pequenos répteis que viveu durante o Período Triássico, os lagerpetídeos.
A origem dos pterossauros sempre foi um daqueles problemas cabeludos da Paleontologia. Os pterossauros, para quem não conhece ou não se recorda, são aqueles répteis voadores com asas membranosas, que viveram durante a chamada “Era dos Dinossauros”, intervalo do tempo que se estende de 251 à 66 milhões de anos atrás.
O problema com os pterossauros é que seus fósseis mais antigos (que tem cerca de 208 milhões de anos), já apresentam características tão diferenciadas, que é difícil traçar a origem evolutiva do grupo. Eles já apresentam, por exemplo, dedos das mãos hiper-alongados, crânio super modificado e diversas outras características derivadas muito bem adaptadas para o voo. A comparação com outros grupos de répteis que viveram no mesmo período é bastante difícil, pois não temos registros de fósseis de organismos com características intermediárias. Logo, descobrir onde o grupo se encaixa na árvore da vida tornou-se um desafio.
Diversos grupos independentes de cientistas têm se debruçado sobre a questão ao longo do tempo. Os resultados específicos são bastante contrastantes, mas em uma coisa, pelo menos, temos concordado, eles pertencem a um grupo chamado de Archosauromorpha, que inclui, por exemplo, os dinossauros e os crocodilos atuais. É um grupo muito amplo. Isso serve para você entender a gravidade da questão.
Há muita discordância sobre em que ponto exato dentro dos Archosauromorpha se encaixam os pterossauros. Alguns pesquisadores sugerem que eles teriam derivado de Archosauromorpha basais. Outros, que eles são parentes bem mais próximos dos dinossauros, derivados de avemetatarsálios basais próximo aos lagerpetídeos. Aí entra o estudo publicado hoje.
Arte de Rodolfo Nogueira.
O estudo publicado hoje na revista Nature é assinado por 18 autores, entre eles os brasileiros Mário Bronzati, Sergio F. Cabreira, Lúcio Roberto da Silva e Max Langer. O que os 18 pesquisadores fizeram foi investigar mais a fundo um grupo específico de pequenos répteis arcossauromorfos do Período Triássico (cerca de 237 a 210 milhões de anos atrás), conhecidos como lagerpetídeos. A equipe analisou não apenas detalhes da aparência externa do esqueleto desses organismos, como utilizou também a tecnologia de tomografia computadorizada para observar dentro de seus ossos. Mais especificamente, dentro do crânio.
Os lagerpetídeos viveram onde hoje é a América do Sul, América do Norte, África e Madagascar. Durante o Triássico, estas massas de terra estavam unidas, formando o supercontinente Pangea. Aqui no Brasil, fósseis de lagerpetídeos são encontrados apenas no Rio Grande do Sul, em rochas da Bacia do Paraná.
Um exemplo de lagerpetídeo brasileiro é Ixalerpeton polesinensis, um animal com cerca de 40 cm de comprimento e 15 centímetros de altura, descrito em 2016. Seu fóssil foi encontrado no município de São João do Polêsine, RS. Ixalerpeton é uma das espécies de lagerpetídeos incluída no estudo. Ele tem preservado parte de sua caixa craniana, o que permitiu aos pesquisadores reconstruir o cérebro e ouvido interno do animal.
Fóssil de Ixalerpeton polesinensis (Foto fornecida pelos autores do estudo)
Outras espécies de lagerpetídeos foram analizadas, como Lagerpeton chanarensis, da Argentina, Dromomeron romeri e Dromomeron gregorii, dos E.U.A. e Kongonaphon kely, de Madagascar. Até pouco tempo atrás, lagerpetídeos eram conhecidos apenas com base em ossos das pernas e do quadril (que se preservam mais facilmente nesse grupo), mas descobertas mais recentes têm revelado mais detalhes sobre a anatomia desses animais, como a aparência do crânio, coluna e membros anteriores.
Os pesquisadores reuniram, então, esses fósseis e fizeram um estudo anatômico comparativo, avaliando diversas características destes animais e comparando-as com de outros Arcosauromorpha. A análise filogenética resultante do estudo (uma forma matemática de análise de parentesco) revelou grandes semelhanças anatômicas entre os lagerpetídeos e os primeiros pterossauros. Apontou, inclusive, a existência de um grupo independente, que incluiria ambas as linhagens, chamado de Pterosauromorpha.
Filogenia calibrada no tempo para Archosauria (Ezcurra et al. 2020).
No trabalho são destacadas várias características únicas compartilhadas apenas por pterossauros e lagerpetídeos, incluindo, por exemplo, alongamento dos ossos do antebraço e das mãos. O parentesco muito próximo surpreendeu bastante, pois anteriormente pensava-se que lagerpetídeos deveriam ser mais aparentados aos silesaurídeos e dinossauros. Assim, o atual estudo estabelece um marco importante para o entendimento da origem dos pterossauros.
Outro resultado fascinante do estudo publicado hoje, envolve a biologia dos lagerpetídeos. Algumas características anatômicas indicam adaptações a um estilo de vida bastante ágil. As análises com tomografia computadorizada e reconstrução do cérebro e ouvido interno desses animais, por exemplo, demonstraram que eles tinham sistemas sensoriais típicos de organismos capazes de movimentos rápidos da cabeça e bom controle do movimento dos olhos e do pescoço. Adaptações como essa são observadas em organismos voadores e/ou arborícolas da atualidade. Talvez os lagerpetídeos utilizassem essa sua característica para a captura de presas ágeis, como insetos. O que é reforçado pela anatomia dos seus dentes.
Dentário (osso anterior da mandíbula) direito do lagerpetídeo Ixalerpeton polesinensis do Triássico do Brasil. Foto fornecida pelos autores do estudo.
Se comparado ao sistema sensorial dos pterossauros, lagerpetídeos tinham basicamente as mesmas adaptações no cérebro e ouvido interno. Isso pode sugerir que características sensoriais vantajosas para o vooevoluíram antes mesmo do voo em sinos Pterosauromorpha (uma pré-adaptação).
Outro ponto interessante é o formato das garras das mãos dos lagerpetídeos, bastante curvadas, o que indica uma função diferenciada dos membros anteriores. Os autores sugerem que elas podem ter sido selecionadas devido a um estilo de vida arborícola (para ajudar a escalar) e/ou utilizadas para a aquisição de presas. O que é interessante , pois pode sugerir um cenário em que o voo nos pterossauros teria evoluído como uma vantagem para se mover de uma árvore para a outra.
Reconstituição em vida do lagerpetídeo Ixalerpeton polesinensis, do Triássico do Rio Grande do Sul, com estilo de vida arborícola. Arte de Rodolfo Nogueira.
Embora este estudo tenha demonstrado um parentesco entre lagerpetídeos e pterossauros, muitas questões ainda permanecem. Entre elas, talvez a mais perturbadora ainda seja como a principal característica dos pterossauros, as asas, evoluíram. Pode parecer frustrante não ter todas as respostas de uma vez, eu sei. Mas estamos chegando cada vez mais perto. Este estudo nos aproxima um pouco mais da “solução” do grande enigma. Pense pelo lado bom: pelo menos, agora sabemos melhor onde procurar respostas!
Nesta sexta-feira, dia 11/12, às 19h, vamos bater um papo ao vivo no nosso canal do Youtube com um dos autores do estudo. Ative o lembrete para não perder:
Pesquisadores brasileiros em parceria com colegas australianos descreveram recentemente, o fóssil de uma espécie de “cobra-cega”, que viveu no Sudeste do Brasil há mais de 85 milhões de anos. Além de o fóssil ser muito importante para o entendimento da evolução do grupo, a espécie é a maior já encontrada entre as cobras-cegas vivas ou extintas. Boipeba tayasuensis, como foi batizada, tinha cerca de 1 metro de comprimento, e sua descoberta preenche uma grande lacuna na história evolutiva das serpentes Scolecophidia.
Boipeba tayasuensis, uma grande cobra-cega do Cretáceo do Brasil. Arte de Jorge Blanco.
O fóssil de Boipeba foi encontrado no município de Taiaçu no Oeste do Estado de São Paulo, próximo à Monte Alto, localidade já conhecida pela ampla ocorrência de fósseis do PeríodoCretáceo. O principal responsável pelo estudo foi Thiago S. Fachini, estudante de doutorado, orientado pela Professora Annie S. Hsiou, ambos da USP de Ribeirão Preto. O estudo ainda contou com a participação de outros dois colegas brasileiros, Silvio Onary e Mário Bronzati, e dois pesquisadores australianos.
O trabalho foi publicado dia 19 de novembro na revista iScience e baseia-se na descrição de uma vértebra bem distinta, grande, para uma “cobra-cega”, e com um formato notavelmente achatado. Daí o nome Boipeba, que significa “cobra-achatada” em Tupi-Guarani. O epíteto específico, “tayasuensis”, faz referência ao município de Taiaçu, assim, a combinação do nome da nova espécie fóssil significa “cobra-achatada de Taiaçu”.
A distinta vértebra de Boipeba tayasuensis (Fachini et al. 2020).
Boipeba tayasuensis era uma serpente de médio porte, com comprimento estimado em 1 metro, tamanho bastante semelhante ao de outras cobras fósseis do mesmo período. O interessante, todavia, é o fato de Boipeba ser uma serpente Scolecophidia, ou seja, um tipo de “cobra-cega”. Atualmente, as cobras-cegas são pequenas serpentes escavadoras, de hábitos essencialmente subterrâneos, que tem os seus olhos bastante reduzidos. As espécies atuais de Scolecophidia não ultrapassam 30 cm de comprimento, o que torna Boipebauma gigante das cobras-cegas. Mesmo as outras formas fósseis conhecidas não têm tamanho comparável ao da “cobra-achatada de Taiaçu”. O fato de ela ser tão grande dá uma pista aos pesquisadores sobre as tendências evolutivas do grupo. A “miniaturização” em Scolecophidia pode ter sido uma tendência mais recente, acompanhando fatores ambientais e ecológicos.
Mas não é só o tamanho que torna Boipeba importante. Fósseis de serpentes são muito raros no Cretáceo, ainda mais na Bacia Bauru, unidade geológica na qual ela foi encontrada. Outros fósseis associados à serpentes já haviam sido descobertos, mas este é o primeiro descrito formalmente como espécie. Boipeba, portanto, amplia o nosso conhecimento sobre a diversidade de organismos do Cretáceo da Bacia Bauru e torna a rede ecológica deste antigo paleoambiente mais complexa.
No Cretáceo brasileiro, o único outro registro inequívoco de uma espécie de serpente é de Seismophis septentrionalis, do Cenomaniano do Maranhão (Bacia de São Luís-Grajaú). Tetrapodophis amplectus, comumente referida como a “cobra com patas” do Aptiano-Albiano da Bacia do Araripe, é questionada por muitos autores e tem uma história bastante complexa (leia mais sobre isso aqui).
Outro aspecto que destaca a descoberta de Boipeba para a Ciência é a idade do seu fóssil. Ela é a espécie mais antiga de cobra-cega já descoberta. Os registros mais antigos de Scolecophidia até então encontrados, eram datados do final do Paleoceno e início do Eoceno da Europa e África (cerca de 56 milhões de anos atrás). Contudo, análises moleculares estimavam o surgimento do grupo para o Cretáceo. Boipeba confirma essa hipótese. A diversificação inicial das cobras-cegas pode ter acontecido na América do Sul e o Brasil pode ter sido um dos palcos principais deste evento evolutivo.
Boipeba estende o registro de Scolecophidia para o Cretáceo Superior do Brasil, preenchendo uma lacuna no espaço e no tempo para a compreensão evolutiva do grupo. As previsões moleculares agora ganharam sustento de evidências paleontológicas.
Boipeba é mais uma descoberta recente que demonstra como o território brasileiro é importante para a Paleontologia mundial. As contribuições que o artigo de Boipeba traz são fundamentais para os estudiosos de evolução de serpentes e, com certeza, atrairão a atenção de paleontólogos do mundo para os estratos rochosos do interior de São Paulo.
NOTA: o grupo mais popularmente conhecido como “cobra-cega” são as cecílias, ou gimnofionas, que são um tipo de anfíbio. As Scolecophidia, um grupo de serpente, todavia, também podem ser chamadas de “cobras-cegas” por conta de seus olhos reduzidos.
Referência:
Fachini, T. S., Onary, S., Palci, A., Lee, M. S. Y., Bronzati, M., Hsiou, A. S. CRETACEOUS BLINDSNAKE FROM BRAZIL FILLS MAJOR GAP IN SNAKE EVOLUTION. iScience, 1-40. doi: https://doi.org/10.101 /j.isci.2020.101834
Uma nova e interessante proposta para explicar as relações evolutivas dos dinossauros e principalmente, a origem dos dinossauros ornitísquios, grupo que inclui o Triceratops e os famosos dinossauros “bico de pato”, acaba de ser apresentada por dois pesquisadores brasileiros da Universidade Federal de Santa Maria (UFSM), RS, Rodrigo Temp Müller e Maurício Garcia. O estudo foi publicado na revista científica Biology Letters e agita ainda mais a discussão sobre as relações evolutivas dos dinossauros.
Arte de Márcio Castro
Não faz muito tempo que um estudo liderado por um pesquisador britânico chamado Matthew Baron, sacudiu a comunidade paleontológica e abriu espaço para novas discussões sobre uma questão fundamental no estudo de dinossauros: a relação evolutiva entre os grandes grupos desses animais.
Em 2017, Baron e colaboradores ressuscitaram uma antiga hipótese sobre a relação dos dinossauros, que propunha que o grupo que inclui os dinossauros carnívoros, como o Tyrannosaurus, era mais proximamente relacionado ao grupo dos dinossauros ornitísquios, aquele que reune Triceratops, Stegosaurus e os chamados “dinos bico de pato”. A hipótese de Baron e colegas veio com algumas novidades, mas já havia sido considerada no passado, tendo sido descartada por diversos estudos em detrimento da proposta mais clássica, que reúne dinossauros carnívoros e os grandes dinossauros “pescoçudos ” em um mesmo grupo chamado Saurischia.
O agito causado por esse artigo foi tanto, que, na época, muito se falou na mídia popular, inclusive, como se tudo que sabíamos sobre dinossauros tivesse que ser reescrito. Pouco tempo passou, todavia, até a publicação de um outro estudo, rebatendo a proposta de Baron e colaboradores. Estudo esse, liderado por um pesquisador brasileiro, da USP de Ribeirão Preto, Prof. Max Langer. O estudo liderado por Langer apontava inconsistências no trabalho de Baron e concluía que a hipótese clássica da divisão dos dinossauros ainda se sustentava com prioridade, apesar de a proposta de Baron não poder ser totalmente descartada, já que era somente “um pouco menos provável que a hipótese tradicional”. Leia mais sobre isso aqui.
A história é longa e a disputa continuou em alguns estudos subsequentes, mas aonde eu quero chegar é que, toda essa discussão ressaltou novamente uma verdade muito incômoda para os estudiosos de dinossauros: nós conhecemos muito pouco sobre os primeiros ornitísquios e não conseguimos dizer com certeza como eles se relacionam com os outros dinossauros. E é aí, finalmente, que o trabalho recém-publicado pelos colegas da UFSM pode ajudar.
Os “caçadores de dinossauros” da UFSM
Rodrigo Temp Müller e Maurício Garcia são dois pesquisadores muito privilegiados, pois têm a honra de estar justamente no local que provavelmente foi o berço dos dinossauros, há aproximadamente 230 milhões de anos. Os fósseis de dinossauros mais antigos conhecidos no mundo são encontrados em rochas dessa idade na Argentina, no sul do Brasil e na porção sul do continente africano. Justamente por estarem trabalhando sobre essas rochas, esses pesquisadores têm acesso direto a fósseis fantástico, que contam a história da aurora desse grupo tão fascinante de animais.
Rodrigo Temp Müller e Maurício Garcia
Atualmente, Rodrigo T. Müller e Maurício Garcia, junto a outros pesquisadores, trabalham vinculados ao Centro de Apoio à Pesquisa Paleontológica (CAPPA-UFSM), em São João do Polêsine, RS, onde se dedicam a estudar a história dos primeiros dinossauros.
A nova hipótese de Müller e Garcia
Verdade seja dita, aorigem dos dinossauros ornitísquios é um grande mistério. Enquanto os fósseis mais antigos de dinossauros saurísquios datam de estratos do meio do Período Triássico, entre 245 e 230 milhões de anos, os fósseis mais antigos de ornitísquios são um pouco mais recentes, da transição Triássico-Jurássico, e têm aproximadamente 200 milhões de anos. Explicar esta ocorrência usando a hipótese tradicional sobre da relação dos dinossauros é um pouco constrangedor, pois implica na existência de “linhagens fantasmas” de ornitísquios, que teriam existido entre ‘pouco tempo depois da origem dos dinossauros’, por volta de 240-230 milhões de anos, até cerca de 206 milhões de anos atrás, idade do fóssil mais antigo de ornitísquio conhecido. Resumindo: ornitísquios necessariamente teriam que ter existido entre ~240 e 206 milhões de anos, mas não temos evidências dos fósseis deles.
A nova hipótese proposta por Müller e Garcia solucionaria esse problema de maneira muito elegante, com algo que sempre esteve bem debaixo do nariz dos pesquisadores: os “Silesauridae”.
As criaturas conhecidas como ‘silessaurídeos’ compõe um grupo de organismos extintos normalmente considerados como parentes muito próximos dos dinossauros. De fato, eles e os primeiros dinossauros são extremamente parecidos, tanto em forma, hábitos, quanto em tamanho (imagem).
Fósseis de ‘silessaurídeos’ são encontrados no Brasil, como é o caso de Sacisaurus agudoensis, um pequeno animal, com cerca de 1,5m de comprimento, que viveu há 225 milhões de anos onde hoje é o Rio Grande do Sul.
Reconstituição artística de Sacisaurus agudoensis por Rodolfo Nogueira.
O que Müller e Garcia fizeram em seu estudo foi testar as relações evolutivas dos grandes grupos de dinossauros, incluindo uma ampla amostragem de silessaurídeos (até então tidos como grupo irmão de Dinosauria) e diversas espécies de dinossauros basais. Eles fizeram uma análise filogenética abrangente, que juntou, pela primeira, vez os dados de espécies há tempos conhecidas, com as espécies mais recentemente descritas. Os resultados encontrados por eles foram surpreendentes…
O que uma análise filogenética faz, basicamente, é testar as relações evolutivas dos organismos por meio da comparação de uma extensa matriz de dados sobre eles. Estes dados podem ser genéticos ou morfológicos, ou os dois, por exemplo. No caso dos fósseis, os pesquisadores normalmente usam dados morfológicos (da forma) para realizá-las, pois informações genéticas só ficam preservadas em materiais relativamente recentes (de até algumas centenas de milhares de anos). O produto de uma análise filogenética são um conjunto de “árvores evolutivas” possíveis, mostrando todas as relações prováveis entre os organismos analisados. Algumas árvores são estatisticamente mais plausíveis que outras e, normalmente são essas as consideradas mais atentamente pelos pesquisadores.
Uma incrível descoberta
O que Müller e Garcia recuperaram em suas análises foi algo diferente de tudo antes proposto e muito excitante para os estudiosos de dinossauros, já que tem o potencial de explicar várias questões sobre o início da história evolutiva do grupo. Em especial, o tal mistério das “linhagens fantasmas” de ornitísquios.
De acordo com os resultados do estudo, os chamados ‘silessaurídeos’ seriam, na verdade, uma série de espécies ordenadas sucessivamente na base de Ornithischia. Ou seja, linhagens que teriam gradualmente acumulado características típicas de ornitísquios ao longo de milhões de anos. Essa hipótese preencheria o intervalo evolutivo antes “assombrado” pela ideia de “linhagens fantasmas”.
Os “silessaurídeos” seriam os dinossauros ornitísquios basais “que faltavam”! De acordo com a hipótese recuperada na análise de Müller e Garcia (2020), ‘Silesauridae’ seriam um grupo parafilético, na base de Ornithiscia. Sob esta perspectiva, Sacisaurus voltaria a ser um dinossauro.
Imagem do artigo de Müller e Garcia (2020).
Outros autores já haviam apontado uma possível relação entre silessaurídeos e ornitísquios, mas o cenário evolutivo encontrado por Müller e Garcia é inédito. Os ornitísquios clássicos, segundo o novo estudo, teriam evoluído a partir de formas típicas de silessaurídeos por meio de mudanças graduais ao longo do tempo, partindo inclusive, de uma possível forma carnívora. O que assemelharia o cenário evolutivo dos ornitísquios ao dos saurísquios sauropodomorfos (veja este vídeo aqui para entender).
Todos os ornitísquios conhecidos até o momento apresentam características que os vinculam a uma dieta herbívora, todavia os mais antigos ‘silessaurídeos’ tinham dentes pontiagudos e afiados, possivelmente adaptados para uma dieta carnívora.
Os pesquisadores responsáveis pela pesquisa reconhecem que ainda é cedo para que a questão relacionada a origem dos dinossauros ornitísquios possa ser considerada totalmente respondida. A nova hipótese de Müller e Garcia agora deverá continuar sendo testada à medida que novos fósseis foram descobertos e novas interpretações realizadas.
Nós aqui também estamos de olho.
A nova proposta de Müller e Garcia (2020). Arte de Márcio Castro.Como ficaria a configuração da nova árvore evolutiva dos dinossauros de acordo com o estudo de Müller e Garcia (2020). Arte de Márcio Castro.
Se você se interessou pelo estudo, você pode ler ele na íntegra AQUI.
Leia também a postagem no blog do parceiro ‘Coelho Pré-Cambriano’: AQUI.
Referência:
Müller, R.T. & Garcia, M. 2020. A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biology Letters, https://doi.org/10.1098/rsbl.2020.0417
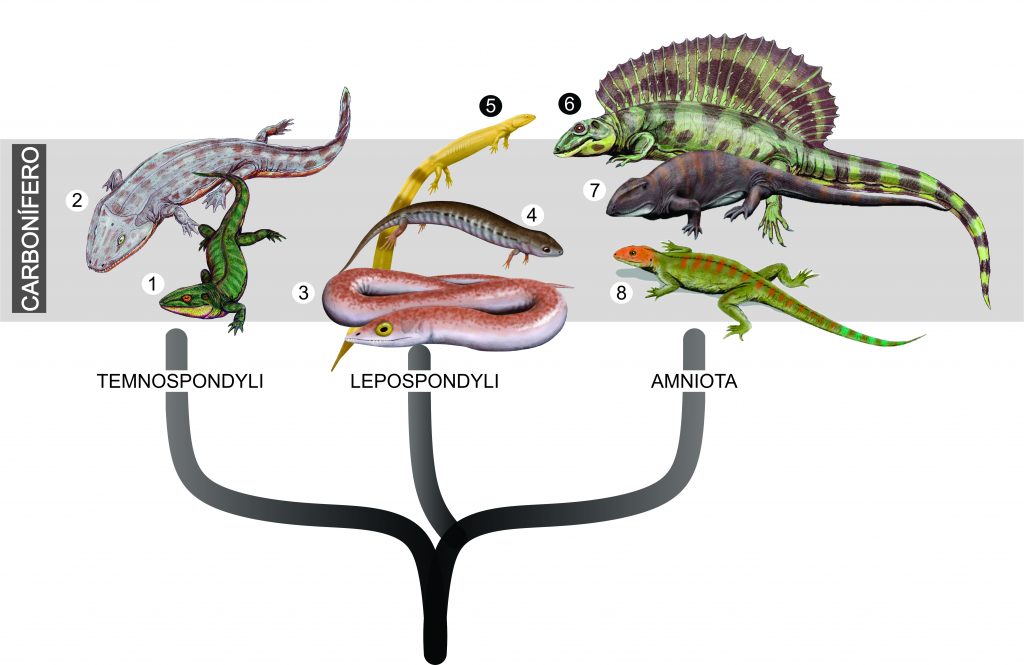
Bem no início da história evolutiva dos tetrápodes, inúmeros animais eram adaptados para a sobrevivência no ambiente aquático. Os ditos anfíbios, na verdade, compunham uma grande diversidade de animais (ex. temnospôndilos, lepospôndilos – Fig. 1), que aterrorizavam áreas inundadas e outros corpos d’água. No entanto, nesse ambiente hostil, surgiram pequenos e frágeis tetrápodes com uma especialização reprodutiva inovadora: o ovo amniótico.
Figura 1. Grupos de tetrápodes mais representativos do Carbonífero. 1. Dendrerpeton. 2. Iberospondylus. 3. Phleghethontia. 4. Hyloplesion. 5. Urocordylus. 6. Ianthasaurus. 7. Ophiacodon. 8. Hylonomus. Ilustrações: 1. ДиБгд; 2; 6. Dmitry Bogdanov; 3-5. Smokeybjb. 7-8. Nobu Tamura (Creative Commons).
Geralmente é ensinado que a presença de membranas diferenciadas e externas ao embrião fizeram com que os tetrápodes conquistassem definitivamente o ambiente terrestre e se diversificassem em formas gigantescas e até mesmo voadoras. Dessa forma, os Amniota ganharam o título de colonizadores do ambiente terrestre. Mas, e o ambiente aquático? Ficou para os anfíbios? Os amniotas deram as costas para o ambiente aquático?
A resposta é NÃO e no Brasil temos um dos grupos fósseis mais importantes para contar essa história: os mesossauros!
Mesossauros?
Talvez os Mesosauridae sejam os amniotas melhor representados e conhecidos no registro fossilífero do nosso país. Seus fósseis são abundantes em rochas permianas da Formação Irati e sua ampla distribuição geográfica (de Goiás até o Rio Grande do Sul) garantiu um bom conhecimento acerca do seu modo de vida e aspectos osteológicos, principalmente da região pós-craniana (Modesto, 2009; Sedor & Ferigolo, 2001).
Os mesossauros fazem parte de um grupo de amniotas conhecido como Parareptilia. Os “pararrépteis” formam uma pequena linhagem que surgiu no Carbonífero e apresenta uma grande diversidade de formas com especializações variadas e novidades evolutivas. Para vocês terem ideia da importância do grupo, nos pararrépteis encontramos: formas herbívoras de grande porte (como os pareiassauros); o mais antigo réptil com capacidade de locomoção bípede conhecido (Eudibamus cursoris); os primeiros amniotas com implantação dentária do tipo tecodonte (dentes implantados em alvéolos, e.g. Bolosaurus grandis) e o primeiro grupo de amniotas a apresentar especializações para o ambiente aquático: sim, os mesossauros.
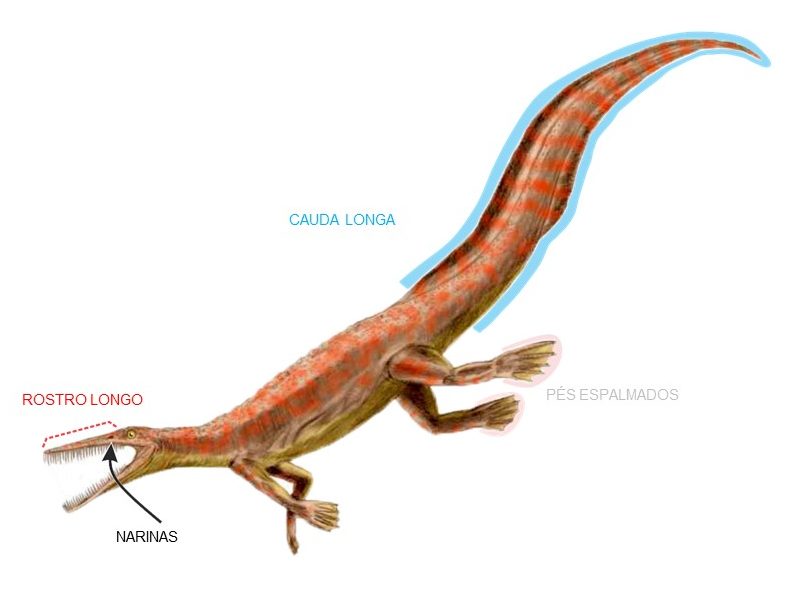
As características que indicam que esses animais eram adaptados para a vida no meio aquático são bastante evidentes (Fig. 2): eles apresentam crânio com rostro alongado (i.e. focinho comprido), narinas próximas às órbitas, coluna vertebral longa, principalmente na região caudal, pés espalmados e ossos com paquiostose e osteosclerose (explico a seguir).
Figura 2. Representação artística de Mesosaurus tenuidens. Ilustração por Nobu Tamura (Creative Commons).
Os mesossauros viviam em um mar extenso (mar Irati-Whitehill) confinado, que cobria áreas que atualmente correspondem ao Brasil, África do Sul, Paraguai e Uruguai (Oelofsen & Araújo, 1983). Nesse grande corpo d’água, os mesossauros compartilhavam o ambiente com… bom, com nenhum outro tetrápode! É isso mesmo! Até o momento, esses pararrépteis são os únicos tetrápodes conhecidos para a Formação Irati. Curioso né?
Na unidade, também ocorrem outros vertebrados (e.g. Chahud & Petri, 2009; 2010), tais como fósseis de peixes (tubarões, celacantos e paleonisciformes) e um dente de um provável anfíbio, mas em camadas de rocha distintas, tornando muito difícil inferir que esses vertebrados compartilhavam o mesmo ambiente numa mesma época.
Se eles não compartilhavam o ambiente com outros vertebrados, então os dentes compridos dos mesossauors eram usados para que?
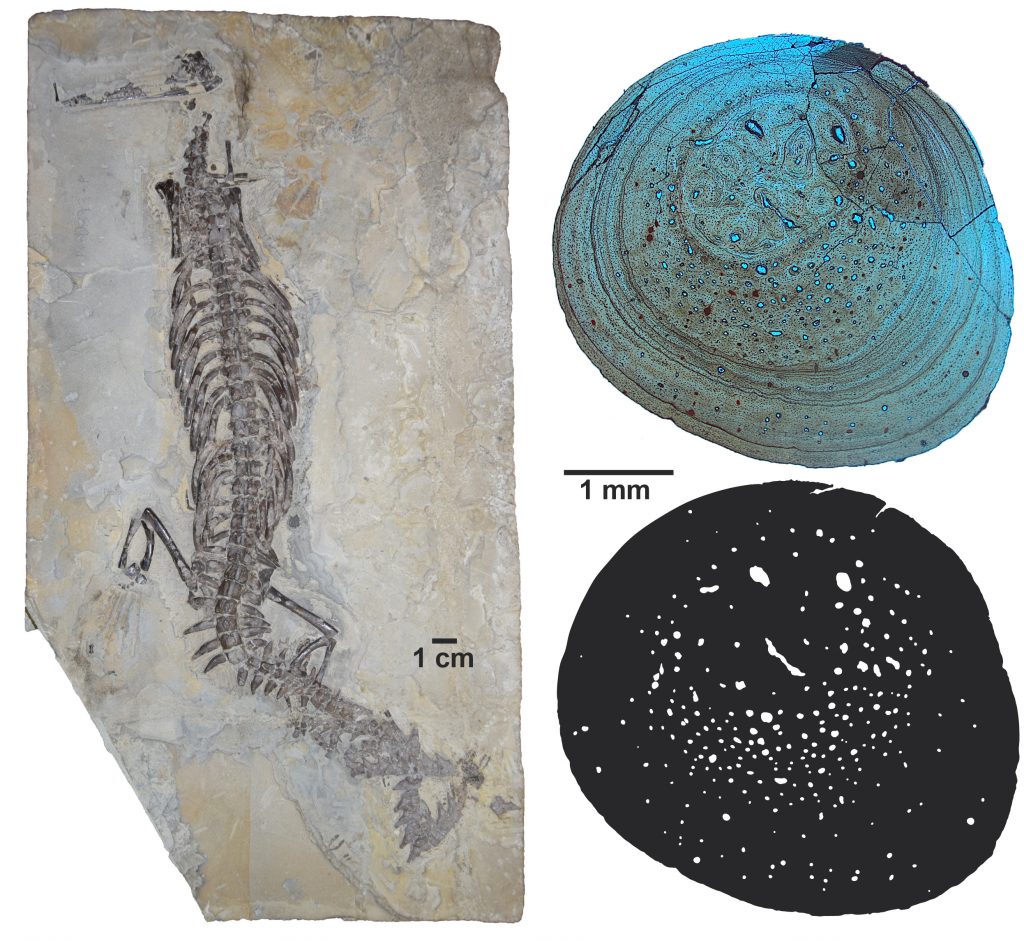
Entre os mesossaurídeos são reconhecidas três espécies (Fig. 3): Brazilosaurus sanpauloensis, Stereosternum tumidum e Mesosaurus tenuidens. Nem todos os mesossauros apresentavam dentes compridos. B. sanpauloensis, por exemplo, apresenta os dentes mais curtos entre os Mesosauridae, mostrando que haveria uma certa variação de dieta entre eles. Sobre a alimentação dos mesossauros, temos fortes pistas: fósseis de crustáceos são encontrados em abundância nas mesmas rochas que nossos pararépteis. Dessa forma, é inferido que pelo menos M. tenuidens se alimentasse desses crustáceos, sendo os dentes utilizados como uma espécie de filtro, que prenderia os invertebrados. Além disso, alguns pesquisadores também sugerem canibalismo e necrofagia, devido a presença de pequenos ossos encontrados em conteúdos gástricos e coprólitos associados de mesossaurídeos (Silva et al. 2017).
Figura 3.A. Stereosternum tumidum. B. Brazilosaurus sanpauloensis. C. Mesosaurus tenuidens.Fotografias: A, Ghedoghedo (Creative Commons); B e C, Thiago Carlisbino – espécimes do Museu de Ciências Naturais, Setor de Ciências Biologicas, UFPR.
Voltando ao assunto dos ossos: Paquiostose? Osteosclerose? O que é isso?
O osso é um tecido vascularizado, composto por células e uma matriz extracelular mineralizada. É um tecido vivo, que tem a função de suporte, proteção, produção de células sanguíneas e também atua como regulador do equilíbrio mineral do organismo (Hall, 2005).
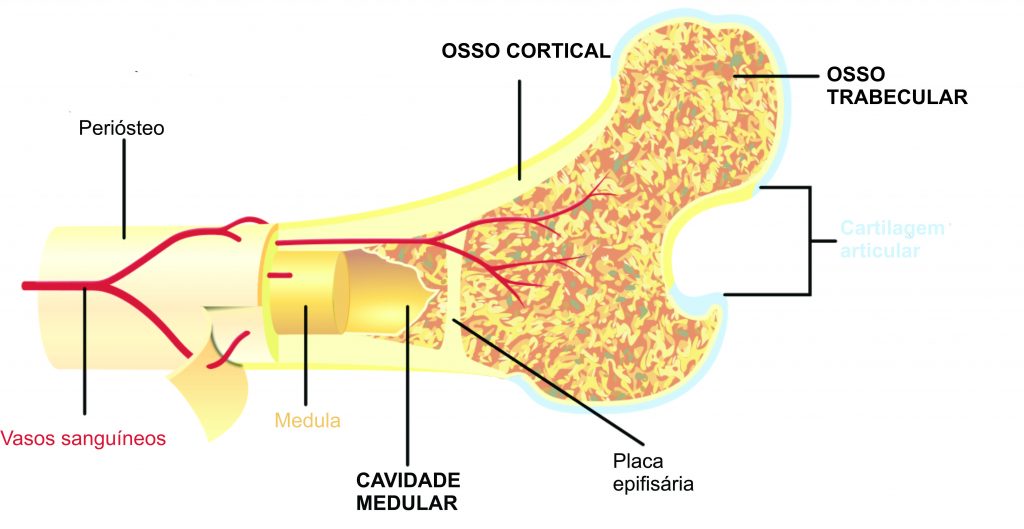
Se pegarmos um osso longo (ex.: fêmur, úmero ou costela) e fizermos um corte na porção média dele, é bem provável veremos uma parte externa mais compacta e outra interna menos mineralizada, mais porosa (variações existem, é claro). A porção periférica é chamada de córtex (osso cortical na imagem), enquanto a interna é chamada de cavidade medular (que contém a medula óssea) (Fig. 4). Em estudos microanatômicos, são detalhadas as variações dessas regiões nos ossos, tais como a espessura do córtex, diâmetro do canal medular e porosidade. O aspecto microanatômico do osso é importante para entender a paleobiologia do vertebrado fóssil, pois sua organização está intimamente relacionada com o hábitat, modo de vida e aspectos biomecânicos do organismo (Krillot et al. 2008). Portanto, é uma ferramenta útil para quem deseja entender como determinado animal vivia em um passado distante.
Figura 4. Representação esquemática de fêmur humano. Ilustração por Pbroks13 (Wikimedia Commons).
Para exemplificar, vou tentar caracterizar os aspectos microanatômicos de ossos longos em espécies terrestres:
Nestas formas, a relação entre espessura cortical, diâmetro do canal medular e seu preenchimento ou não por trabéculas ósseas, parece depender muito do porte do animal.
No geral, amniotas terrestres apresentam ossos dos membros com córtex delgado, canal medular amplo e sem a presença de trabéculas. No entanto, a situação muda quando o animal se torna muito grande. Imaginem um osso extremamente comprido e com poucos milímetros de espessura cortical: será que ele aguentaria suportar toneladas e ao mesmo tempo resistir ao estresse mecânico provocado pela locomoção? É bem provável que não. Dessa forma, é comum que o osso desses animais apresente córtex espesso e cavidade medular preenchida por osso trabecular (i.e. em forma de travas) (Fig. 5).
Figura 5. Organização microanatômica de ossos longos de A. Canis lupus (terrestre), B. Chelus fimbriatus (habita águas rasas), C. Rhinocerus unicornis (terrestre de grande porte). Reparem na vascularização e no preenchimento do canal medular (porção mais ao centro da ilustração). Ilustrações dos cortes transversais: A-B, retirado de Krillof et al. (2008); C, retirado de Houssaye et al. (2015).
Já a vida no meio aquático impõe uma série de restrições mecânicas diferenciadas, devido à água apresentar maior densidade e viscosidade que o ar. Nos ossos é comum o aumento em massa e densidade, que ocorre devido à grande quantidade de deposição de osso cortical e o aumento da compacidade interna. Dessa forma, ossos paquiostóticos possuem aspecto inchado, devido à grande quantidade de osso cortical depositado, enquanto que ossos osteoscleróticos são extremamente compactos, quase não apresentando canais vasculares e, quando apresentam, são geralmente de pequeno calibre. Nos mesossaurídeos, tanto a paquiostose, quanto a osteosclerose ocorrem em vértebras e costelas (Fig. 6). Tal característica é denominada de paquiosteosclerose.
Figura 6.A. Stereosternum tumidum (espécime pertencente à Coleção do Laboratório de Paleontologia de Vertebrados da UFRGS). B. Secção transversal de costela de S. tumidum. Reparem no tamanho do canal medular em relação ao osso cortical. C. Ilustração esquemática que ressalta o baixo grau de porosidade na costela de S. tumidum. Fotografias e ilustração por Thiago Carlisbino.
No entanto, essa adaptação tem um preço: o aumento da massa esquelética tem consequências na locomoção subaquática (Taylor, 2000). A paquiostose parece ocorrer com mais frequência em espécies que habitam áreas rasas aonde o animal tem a possibilidade de alcançar o fundo com certa facilidade. Táxons de uma mesma linhagem, mas adaptados ao ambiente pelágico (que vivem em mar aberto) tendem a não apresentar tal característica o que não acontece em formas “menos ágeis” (Houssaye, 2009). De fato, os mesossauros não alcançavam grande velocidade na água e, além disso, os indivíduos encontrados em rochas depositadas em ambiente mais profundo (ex. folhelhos), são geralmente os de maior tamanho (Oelofsen & Araújo, 1983; Villamil et al. 2015).
Além dos aspectos ósseos, há outras evidências que sugerem o habitat preferencial dos mesossauros?
Além dos fósseis de mesossauros ocorrerem apenas em rochas que foram depositadas em ambiente marinho, também temos evidências icnológicas. Sedor & Silva (2004), descreveram pela primeira vez marcas subaquáticas de mesossaurídeos em calcários da Formação Irati no Estado de Goiás. As marcas foram produzidas pelo toque dos pés no sedimento durante o impulso para aumento de velocidade ou mudança de curso. Posteriormente, Silva et al. (2009) estudaram pegadas de mesossauros semelhantes de material oriundo do Estado do Paraná e descreveram a icnoespécie Mesosaurichnium natans (Fig. 7).
Figura 7. Marcas subaquáticas produzidas por mesossaurídeos (Mesosaurichnium natans). Figura retirada de Silva & Sedor (2017).
Apesar do estudo microscópico dos fósseis se dar a partir de métodos destrutivos (lembrem-se de ter muito cuidado na escolha dos espécimes!), ele fornece detalhes muito interessantes acerca da paleobiologia e paleoecologia dos táxons fósseis. Neste texto, descrevi brevemente o que podemos inferir a partir dos detalhes microanatômicos dos ossos. No entanto, existem outros aspectos que podemos interpretar a partir de lâminas delgadas de fósseis: detalhes do crescimento, a taxa de deposição óssea e a possibilidade de se inferir a idade de um indivíduo são apenas algumas delas.
Nos mesossauros a análise da microestrutura óssea ainda está no início e tem muita coisa para ser mostrada.
Até mais!
—————————
Quer saber mais sobre esses fofos? Corre lá no canal pra conferir dois vídeos super bacana que a gente fez sobre eles!
CHAHUD, A., PETRI, S. (2009). Novos Xenacanthidae (Chondrichthyes, Elasmobranchii) da base do Membro Taquaral, Formação Irati, Permiano da Bacia do Paraná. Revista do Instituto Geológico, 30(1-2), 19-24.
CHAHUD, A., PETRI, S. (2010). Anfíbio e Paleonisciformes da Porção Basal do Membro Taquaral, Formação Irati (Permiano), Estado de São Paulo, Brasil. Geologia USP: Série Científica, 10(1), 29-37.
HALL, B. K. (2005). Bones and cartilage: Developmental and and evolutionary skeletal biology. Elsevier, 2005, 792p.
HOUSSAYE, A. (2009). “Pachyostosis” in aquatic amniotes: a review. Integrative Zoology, 4 (4), 325-340.
HOUSSAYE, A. et al. (2015). Biomechanical evolution of solid bones in large animals: a microanatomical investigation. Biological Journal of the Linnean Society, 117, 50-371.
KRILLOF, A. et al. (2008). Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference. Journal of Evolutionary Biology, 21, 807-826.
MODESTO, S. P. (2010). The postcranial skeleton of the aquatic parareptile Mesosaurus tenuidens from the gondwanan Permian. Journal of Vertebrate Paleontology, 30(5), 1378-1395.
OELOFSEN, B. W. & ARAÚJO, D. C. (1983). Palaeoecological implications of the distribution of mesosaurid reptiles in the Permian Irati Sea (Paraná Basin), South America. Revista Brasileira de Geociências, 13, 1-6.
SEDOR, F. A. & FERIGOLO, J. (2001). A coluna vertebral de Brazilosaurus sanpauloensis Shikama & Ozaki, 1966 da Formação Irati, Permiano da Bacia do Paraná (Brasil) (Proganosauria, Mesosauridae). Acta Biologica Paranaense, 30, 151-173.
SEDOR, F. A. & SILVA, R. C. (2004). Primeiro registro de pegadas de Mesosauridae (Amniota, Sauropsida) na Formação Irati (Permiano Superior da Bacia do Paraná) do Estado de Goiás, Brasil. Rev. Bras. Paleontol. 7, 269–274. doi: 10.4072/rbp.2004.2.21
SILVA, R. C., SEDOR, F. A., FERNANDES, A. C. S. (2009). Ichnotaxonomy, functional morphology and paleoenvironmental context of Mesosauridae tracks from Permian of Brazil. Rev. Bras. Geociências. 39, 705–716.
SILVA, R. C. & SEDOR, F. A. (2017). Mesosaurid swim traces. Frontiers in Ecology and Evolution, 5: 22.
SILVA, R. R. et al. (2017). The feeding habits of Mesosauridae. Frontiers in Earth Science, 5: 23.
TAYLOR, M. A. (2000). Functional significance of bone ballast in the evolution of buoyancy control strategies by aquatic tetrapods. Historical Biology 14, 15–31.
VILLAMIL, J. et al. (2015). Optimal swimming speed estimates in the Early Permian mesosaurid Mesosaurus tenuidens (Gervais, 1865) from Uruguay. Historical Biology, 28, 963-971.

























{kind=link}